

Breaking the Code | Ep. 1009
How Enhancers Are Rewriting the Rules of Genetics
Hello Avatar! Welcome back for another week of biotech analysis. Today is Sunday, which means this is our Building Biotech newsletter that is focused on discussing biopharma strategy topics. This week, we dive into one of the most important functional genomics papers of the decade, and its massive implications for biotech. While most of the industry still treats enhancers as black-box regulators, a new enhancer dissection atlas breaks them down block by block, base by base. The result? A blueprint for designing safer gene therapies, fine-tuning epigenetic editors, and decoding previously undruggable disease drivers. In this issue, we unpack what this means for variant interpretation, tissue-selective delivery, and the companies already building pipelines around it.
We are now publishing 7x per week according to the following cadence:
Mondays: Stocks
Tuesdays: Biotech
Wednesdays: Podcast
Thursdays: Markets
Fridays: News
Saturdays: Podcast
Sundays: Strategy
We are also publishing unique content on X - be sure to follow up if you are not already @BowTiedBiotech. And to check-out the archive of our work on X you can find it on our website at: BowtiedBiotech.subtack.com/x-articles.
SUBSCRIBE TO PODCAST HERE:
Please help spread the work by subscribing and hitting the share button if you are enjoying our bi-weekly newsletters!

Enough shilling for the day, lots to cover this week, let's get started!
What Exactly Is an Enhancer?
Enhancers are short, non-coding stretches of DNA that control when, where, and how strongly genes are expressed. They don’t encode proteins. Instead, they act like molecular switches, binding transcription factors and looping into contact with promoters to activate or amplify gene transcription. One gene can be regulated by dozens of enhancers, and a single enhancer can regulate multiple genes depending on cellular context.
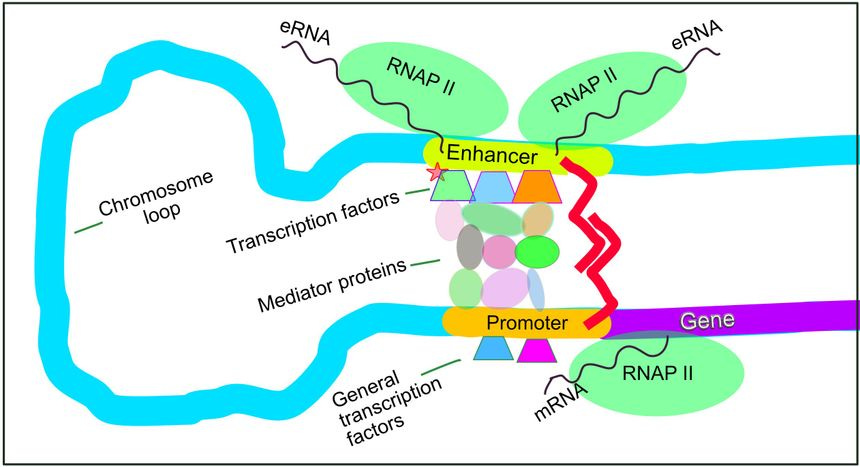
What makes enhancers so powerful is their precision. Unlike promoters, which sit directly upstream of genes, enhancers can be located thousands of base pairs away, upstream, downstream, or even inside introns. Their activity is tightly restricted by tissue type, developmental stage, and chromatin structure. That’s why the same genome can produce a neuron, a liver cell, or a T-cell, because different enhancers are “on” in each context.
To-date the problem hasn’t been finding enhancers, it has been understanding how they worked. Sequencing techniques like ATAC-seq and ChIP-seq showed us which regions of DNA were open and bound by transcription factors, hinting at enhancer activity. But these tools paint with a broad brush. They highlight hundreds or thousands of base pairs, without telling you which specific sequences within that region are actually required for function. It’s like seeing a city from space, you know it’s there, but you can’t tell which buildings keep the lights on.
Without fine-grained resolution, developers couldn’t pinpoint which mutations disrupted enhancer output, or which motifs to use for synthetic control. You couldn’t predict off-target expression, test combinatorial effects, or design gene therapies that relied on consistent enhancer behavior. Therapeutically, that meant enhancers were risky and unprogrammable, useful in theory, but unreliable in practice.
This Isn’t Just Regulatory DNA. It’s a Therapeutic Toolkit.
We’ve spent the last 20 years decoding genes. But the future of drug development might be written in the space between them.
Enhancers (those cryptic, non-coding switches scattered across the genome) have long been recognized as important. But until now, they’ve been too messy, too unpredictable, and too poorly understood to engineer with intent. That just changed.
A recent study from the ENCODE consortium offers a first-of-its-kind map that dissects individual enhancers with single-base precision using high-throughput mutagenesis and transgenic mouse models. Think of it like a Google Maps update, not just roads, but traffic, potholes, and every lane. It’s not just that we can now see where enhancer activity comes from. We can turn it off, turn it up, or redirect it entirely.
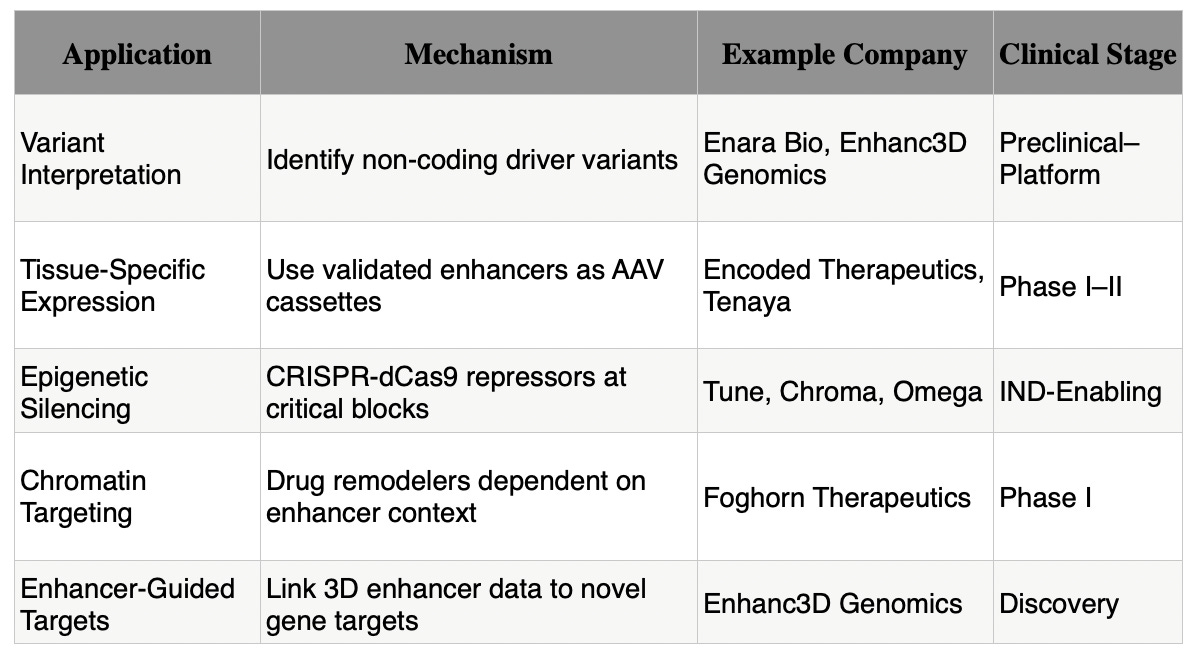
That shift unlocks an entire menu of therapeutic applications. From precision variant interpretation to plug-and-play tissue-specific expression, enhancer biology is now programmable. And unlike traditional CRISPR edits, we don’t have to cut the genome to get results.
Table 1. Therapeutic Applications of Enhancer Biology
Achilles’ Heels and Hidden Switches
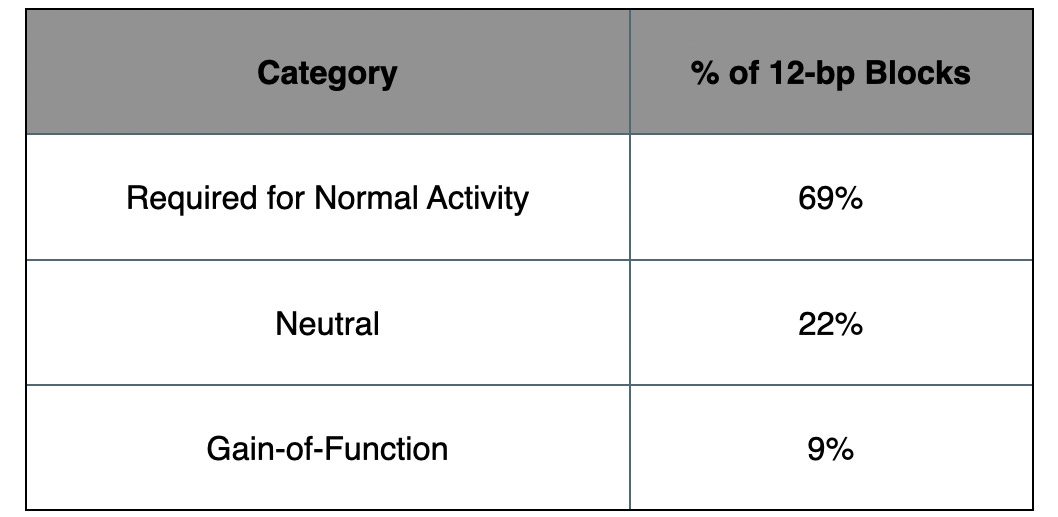
The paper shares an enhancer dissection map which flips the script on what we know to-date. By systematically mutating each 12-bp block and testing in vivo output across tissues, it shows us not just where enhancers are, but what makes them tick, and what makes them fail.
The headline: across seven developmental enhancers, 69% of the 12-bp blocks were required for normal activity. But three had single "Achilles' heel" motifs where one tweak knocked out the whole enhancer. Those motifs aren’t just hotspots, they’re access points. And they can now be targeted, edited, or silenced with precision.
Table 2. Percent of 12-bp Blocks with Functional Impact
This kind of functional granularity is unprecedented. Most previous enhancer models were probabilistic and vague. This study uses a combination of saturation mutagenesis, single-cell transcriptomics, and in vivo enhancer-reporter assays to draw crisp boundaries between “essential,” “nonessential,” and even “gain-of-function” sequences. It’s the first clear demonstration that enhancer output can be modular, predictable, and context-specific.
Importantly, these aren't just theoretical results. In some cases, a single mutation didn’t just drop expression in the target tissue, it turned on expression in completely different tissues like brain or heart. That’s a warning and a feature. Because if you can engineer cell-selective activation, you can also unintentionally cause cross-talk. The only way around that? Whole-organism, multi-tissue validation. And that’s exactly what this team did.
The New Logic of Variant Interpretation
The clinical implications are immediate. The next time a geneticist sees a non-coding variant of uncertain significance (VUS) near a developmental gene, they no longer have to shrug. This enhancer dissection map gives them a yes/no answer at the block level. The same applies for rare disease researchers hunting elusive causes of congenital disorders.
By tying single variants to sharp drops or gains in enhancer activity, this work lays the foundation for VUS triage based on functional impact rather than guesswork. That’s a game-changer for both diagnostics and enrollment. With the right enhancer maps, we can reclassify inherited risk variants and match patients to enhancer-modulating therapies in oncology, immunology, and neurodevelopment.
Companies like Enara Bio are already leaning into this approach. Their “dark antigen” platform sources tumor-specific enhancers that get dysregulated by non-coding mutations, producing neoantigens with minimal normal tissue expression. That’s ideal for TCR-based cell therapies and vaccines. With data from this study, they can now prioritize the highest-impact enhancer blocks to amplify expression before priming the immune system.

Gene Therapy’s New Safety Valve
Traditional gene therapy carries an unavoidable tradeoff: potency versus control. Once you deliver a gene, it's hard to modulate its expression across different tissues. That’s why enhancer-guided tissue targeting is such a breakthrough. Rather than using ubiquitous promoters or random regulatory regions, we can now borrow validated, cell-selective enhancers with precisely characterized boundaries.
For example, Encoded Therapeutics uses a GABAergic-neuron enhancer to selectively boost SCN1A in inhibitory neurons for Dravet syndrome. That’s not a guess, it’s a synthetic design built from enhancer logic. Their program, ETX101, is already in the clinic. It’s the first of a wave of enhancer-cassette AAVs that combine gene delivery with spatial control. With this new paper, developers gain a library of modular switches they can mix and match.

Tenaya Therapeutics is doing the same in cardiology. Their cardiac gene therapy programs combine capsid engineering with enhancer-guided expression. The safety advantage is obvious: deliver broadly, express locally. And as the study showed, even subtle enhancer changes can toggle expression up or down without introducing off-target effects, if the right motifs are used.
Editing Without Cutting: Enter Epigenetic Control
If you know exactly which base pair activates an enhancer, you don’t have to delete it. You can silence it. That’s where CRISPR-based epigenetic editors come in. These systems, like dCas9 fused to histone-modifying enzymes, let you write over enhancer states like a whiteboard, no double-strand breaks required.
The best targets? The “Achilles’ heel” motifs uncovered in this study. That’s why Tune Therapeutics and Chroma (nChroma) Bio are racing to build enhancer silencing programs. They’re aiming for reversible, tunable control over gene expression, starting in the liver and CNS. Tune’s lead program is expected to enter clinical trials this year. The logic is simple: turn down disease-driving genes by switching off their upstream enhancer hubs.

Omega Therapeutics (now shutdown) takes the concept even further. Their Epigenomic Controllers modulate entire gene expression domains by targeting insulated genomic domains (IGDs), many of which are built from enhancer blocks. The high-resolution maps in this paper make those blocks visible, and editable. That shrinks the distance between enhancer biology and drug-like action.
A Shortcut to Chromatin Drug Targets
Enhancers don’t work in isolation. They depend on chromatin remodelers, looping complexes, and transcription factor cooperation. By identifying which enhancer motifs fail without specific chromatin regulators, this study gives a new readout for small-molecule target discovery.
Foghorn Therapeutics is ahead of the curve here. They’re building synthetic lethal combinations between remodeler mutations (like SMARCA4 loss) and dependency on remaining enhancer machinery. Their lead program, FHD-909, selectively collapses enhancer activity in tumors that lose BRG1 but retain BRM. This paper strengthens that rationale, showing how enhancer architecture depends on local chromatin context, which can now be modeled with resolution.
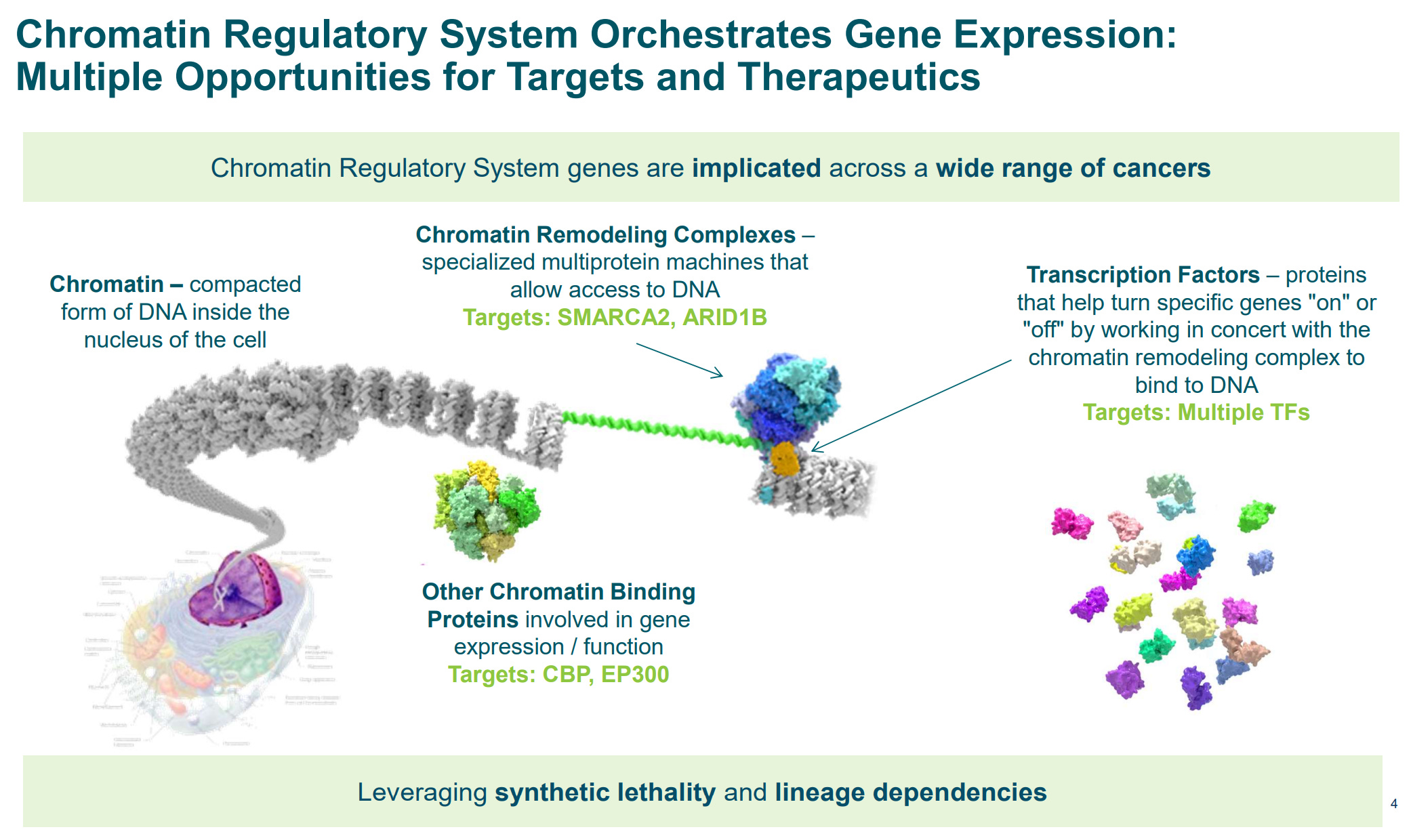
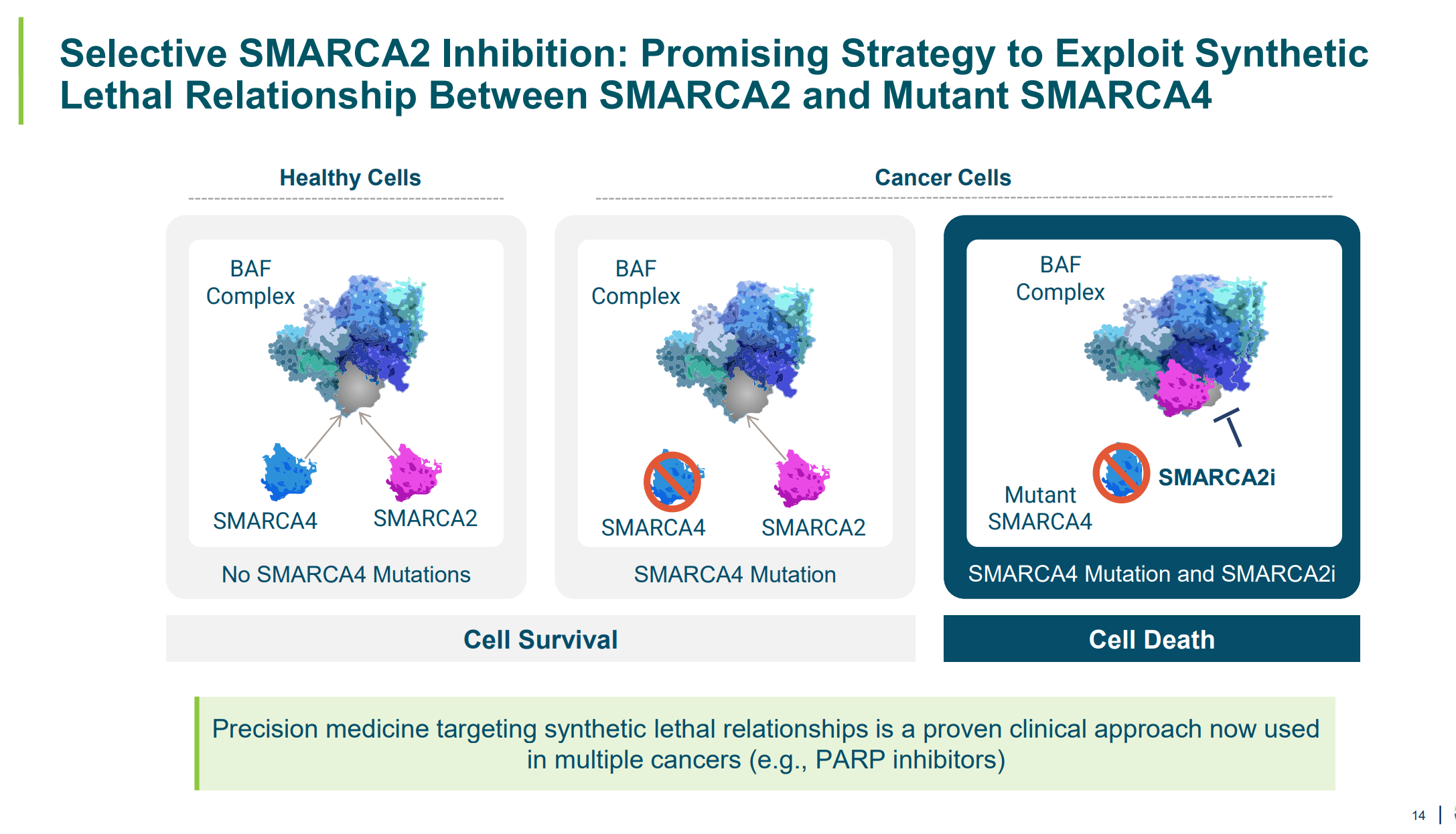
With this level of enhancer annotation, drug hunters can reverse-engineer dependencies. Which enhancers fail when BAF complexes are disrupted? Which enhancer gains are mediated by specific histone marks? These aren’t abstract questions anymore. They’re input for new chemical screens.
From Maps to Models: The New Development Pipeline
Beyond the biology, this study provides a reproducible pipeline for enhancer validation.
Start with an ATAC-seq model like ChromBPNet to predict accessible motifs.
Run mutagenesis on 12-bp tiles.
Plug that into a mouse enhancer-reporter transgene, and run multi-tissue single-cell RNA-seq.
Then integrate expression changes with computational motif impact.
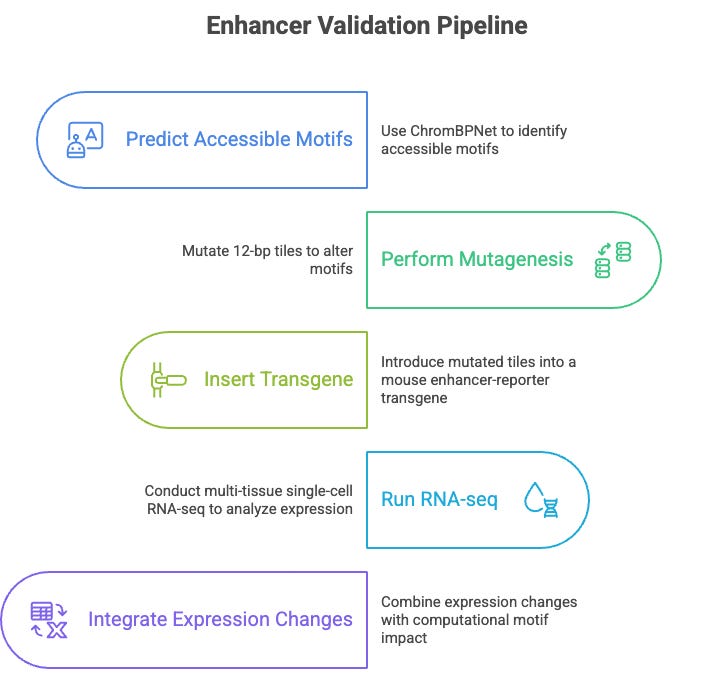
Every step is scalable.
That workflow is already being mirrored by tools like STARR-seq, scMPRA, and high-throughput barcoded reporters. But this paper’s mouse-integration layer adds critical in vivo fidelity, especially for tissue cross-talk. Therapeutic developers should be building in similar validation loops, particularly for programs with narrow safety margins.
And the data are generalizable. These aren’t just one-off enhancers. They span endoderm, mesoderm, and ectoderm lineages. That means developers working in lung, gut, muscle, kidney, skin, and brain all have relevant scaffolds. Instead of screening hundreds of candidates, they can start with a rational subset of pre-validated enhancer tiles.
How to Operationalize It
Start with your gene of interest. What enhancers are known to regulate it? Use ATAC-seq and Hi-C to map candidate regions. Then overlay the enhancer atlas from this paper to see if known sensitivity motifs overlap. If they do, you have a hit. If they don’t, you still have a roadmap to test gain-of-function designs.
Next, decide your therapeutic intent. Are you trying to boost, restrict, or silence expression? If the goal is tissue-specific upregulation, find the strongest enhancer with minimal off-target activity in this map. If you're silencing, aim for the critical 12-bp motifs and dock an epigenetic effector. And if you're building a new construct, splice in one of the validated enhancer tiles as a control module.
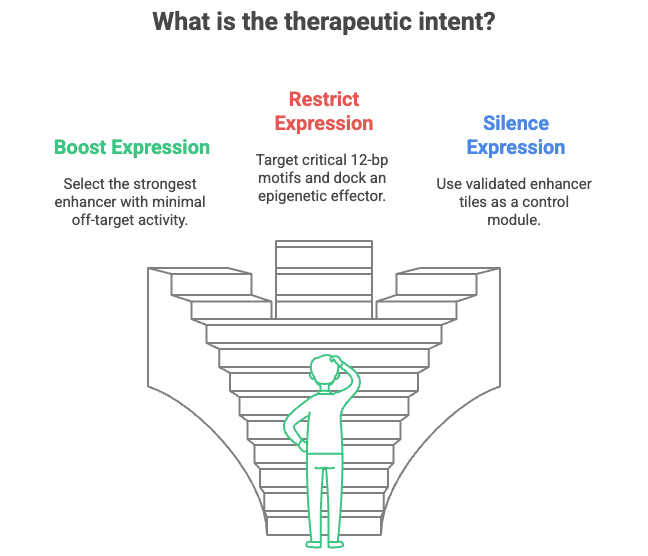
Companies can go further. Build enhancer variant libraries in multiplexed MPRA platforms. Feed that into a dCas9 screening library. Validate best-in-class tiles in mouse models with known tissue expression. Then license or develop programs around enhancer control, either as the therapy or as a gatekeeper for broader cell- or tissue-specific expression.
The Frontier Just Got Closer
This isn’t a vague promise of non-coding DNA utility. This is the moment non-coding biology became legible, targetable, and therapeutically useful. And unlike traditional drug targets, enhancers aren’t proteins with conserved binding pockets. They’re modular regulators of expression. That opens up a whole new axis of control in drug development, one based on when, where, and how genes turn on.

The companies moving fastest here, Enara, Encoded, Tune, Chroma, Omega, Foghorn, are leveraging this shift to control biology at the level of regulatory logic. And as this paper shows, the rules of that logic are now written down. Not as speculation. As experimentally validated maps.
For investors, this marks a new platform category. For scientists, it’s a set of coordinates. For therapeutic developers, it’s the difference between noise and signal. This study doesn’t just open a door. It hands us the key.
CONCLUSION
Today, enhancer biology is no longer an interpretive art. It’s an engineering discipline. With single-base sensitivity maps, predictive motif models, and in vivo validation pipelines, developers can now manipulate non-coding DNA with the same precision they once reserved for exons. The biotech companies that internalize this shift, by licensing the tools, building enhancer-based cassettes, or launching programmable editing programs, are going to have a first-mover advantage in a space most of the world hasn’t even started watching.
As a reminder, if looking to go deeper into the topics we cover check out our website BowTiedBiotech.com, or DM us on twitter, or email us: bowtiedbiotech@gmail.com
DISCLAIMER
None of this is to be deemed legal or financial advice of any kind. All updates are sourced from publicly available disclosures. Insights are *opinions* written by an anonymous cartoon/scientist/investor.
TOP BOWTIEDBIOTECH NEWSLETTERS